Отправить свою хорошую работу в базу знаний просто. Используйте форму, расположенную ниже
Студенты, аспиранты, молодые ученые, использующие базу знаний в своей учебе и работе, будут вам очень благодарны.
Размещено на http://www.allbest.ru/
Мышечные ткани объединяет способность к сокращению.
Особенности строения: сократительный аппарат, занимающий значительную часть в цитоплазме структурных элементов мышечной ткани и состоящий из актиновых и миозиновых филаментов, которые формируют органеллы специального назначения - миофибриллы .
Мышечные ткани представляют собой группу тканей различного происхождения и строения, объединенных на основании общего признака - выраженной сократительной способности, благодаря которой они могут выполнять свою основную функцию - перемещать тело или его части в пространстве.
Важнейшие свойства мышечных тканей. Структурные элементы мышечных тканей (клетки, волокна) обладают удлиненной формой и способны к сокращению благодаря мощному развитию сократительного аппарата. Для последнего характерно высокоупорядоченное расположение актиновых и миозиновых миофиламентов, создающее оптимальные условия для их взаимодействия. Это достигается связью сократимых структур с особыми элементами цитоскелета и плазмолеммой (сарколеммой), выполняющими опорную функцию. В части мышечных тканей миофиламенты образуют органеллы специального значения - миофибриллы. Для мышечного сокращения требуется значительное количество энергии, поэтому в структурных элементах мышечных тканей имеется большое количество митохондрий и трофических включений (липидных капель, гранул гликогена), содержащих субстраты - источники энергии. Поскольку мышечное сокращение протекает с участием ионов кальция, в мышечных клетках и волокнах хорошо развиты структуры, осуществляющие его накопление и выделение - агранулярная эндоплазматическая сеть (саркоплазматическая сеть), кавеолы.
Классификация мышечных тканей основана на признаках их (а) строения и функции (морфофункциональная классификация) и (б) происхождения (гистогенетическая классификация).
Морфофункциональная классификация мышечных тканей выделяет поперечнополосатые (исчерченные) мышечные ткани и гладкую мышечную ткань. Поперечнополосатые мышечные ткани образованы структурными элементами (клетками, волокнами), которые обладают поперечной исчерченностью вследствие особого упорядоченного взаиморасположения в них актиновых и миозиновых миофиламентов. К поперечнополосатым мышечным тканям относят скелетную и сердечную мышечную ткани. Гладкая мышечная ткань состоит из клеток, не обладающих поперечной исчерченностью. Наиболее распространенным видом этой ткани является гладкая мышечная ткань, входящая в состав стенки различных органов (бронхов, желудка, кишки, матки, маточной трубы, мочеточника, мочевого пузыря и сосудов).
Гистогенетическая классификация мышечных тканей выделяет три основных типа мышечных тканей: соматический (скелетная мышечная ткань), целомический (сердечная мышечная ткань) и мезенхимный (гладкая мышечная ткань внутренних органов), а также два дополнительных: миоэпителиальные клетки (видоизмененные эпителиальные сократимые клетки в концевых отделах и мелких выводных протоках некоторых желез) и мионейральные элементы (сократимые клетки нейрального происхождения в радужке глаза).
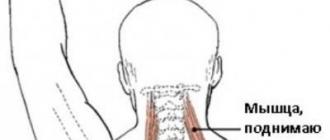
Скелетная поперечнополосатая (исчерченная) мышечная ткань по своей массе превышает любую другую ткань организма и является самой распространенной мышечной тканью тела человека. Она обеспечивает перемещение тела и его частей в пространстве и поддержание позы (входит в состав локомоторного аппарата), образует глазодвигательные мышцы, мышцы стенки полости рта, языка, глотки, гортани. Аналогичное строение имеет нескелетная висцеральная исчерченная мышечная ткань, которая обнаруживается в верхней трети пищевода, входит в состав наружных анального и уретрального сфинктеров.
мышечный миоцит сердечный скелетный
Скелетная поперечнополосатая мышечная ткань развивается в эмбриональном периоде из миотомов сомитов, дающих начало активно делящимся миобластам - клеткам, которые располагаются цепочками и сливаются друг с другом в области концов с образованием мышечных трубочек (миотубул) , превращающихся в мышечные волокна. Такие структуры, образованные единой гигантской цитоплазмой и многочисленными ядрами, в отечественной литературе традиционно именуют симпластами (в данном случае - миосимпластами), однако этот термин отсутствует в принятой международной терминологии. Некоторые миобласты не сливаются с другими, располагаясь на поверхности волокон и давая начало миосателлитоцитам - мелким клеткам, которые являются камбиальными элементами скелетной мышечной ткани. Скелетная мышечная ткань образована собранными в пучки поперечнополосатыми мышечными волокнами , являющимися ее структурно-функциональными единицами.
Мышечные волокна скелетной мышечной ткани представляют собой цилиндрические образования вариабельной длины (от миллиметров до 10-30 см). Их диаметр также широко варьирует в зависимости от принадлежности к определенной мышце и типу, функционального состояния, степени функциональной нагрузки, состояния питания и других факторов. В мышцах мышечные волокна образуют пучки, в которых они лежат параллельно и, деформируя друг друга, часто приобретают неправильную многогранную форму, что особенно хорошо видно на поперечных срезах. Между мышечными волокнами располагаются тонкие прослойки рыхлой волокнистой соединительной ткани, несущие сосуды и нервы - эндомизий. Поперечная исчерченность скелетных мышечных волокон обусловлена чередованием темных анизотропных дисков (полос А) и светлых изотропных дисков (полос I). Каждый изотропный диск рассекается надвое тонкой темной линией Z - телофрагмой . Ядра мышечного волокна - сравнительно светлые, с 1-2 ядрышками, диплоидные, овальные, уплощенные - лежат на его периферии под сарколеммой и располагаются вдоль волокна. Снаружи сарколемма покрыта толстой базальной мембраной, в которую вплетаются ретикулярные волокна.
Миосателлитоциты (клетки-миосателлиты) - мелкие уплощенные клетки, располагающиеся в неглубоких вдавлениях сарколеммы мышечного волокна и покрытые общей базальной мембраной (см. рис. 88). Ядро миосателлитоцита - плотное, относительно крупное, органеллы мелкие и немногочисленные. Эти клетки активируются при повреждении мышечных волокон и обеспечивают их репаративную регенерацию. Сливаясь с остальной частью волокна при усиленной нагрузке, миосателлитоциты участвуют в его гипертрофии.
Миофибриллы образуют сократительный аппарат мышечного волокна, располагаются в саркоплазме по ее длине, занимая центральную часть, и отчетливо выявляются на поперечных срезах волокон в виде мелких точек.
Миофибриллы обладают собственной поперечной исчерченностью, причем в мышечном волокне они располагаются столь упорядоченно, что изотропные и анизотропные диски разных миофибрилл совпадают между собой, обусловливая поперечную исчерченность всего волокна. Каждая миофибрилла образована тысячами повторяющихся последовательно связанных между собой структур - саркомеров.
Саркомер (миомер) является структурно-функциональной единицей миофибриллы и представляет собой ее участок, расположенный между двумя телофрагмами (линиями Z). Он включает анизотропный диск и две половины изотропных дисков - по одной половине с каждой стороны. Саркомер образован упорядоченной системой толстых (миозиновых) и тонких (актиновьх) миофиламентов. Толстые миофиламенты связаны с мезофрагмой (линией М) и сосредоточены в анизотропном диске,
а тонкие миофиламенты прикреплены к телофрагмам (линиям Z), образуют изотропные диски и частично проникают в анизотропный диск между толстыми нитями вплоть до светлой полосы Н в центре анизотропного диска.
В мышечной, как в других тканях, различают два вида регенерации - физиологическую и репаративную. Физиологическая регенерация проявляется в форме гипертрофии мышечных волокон, что выражается в увеличении их толщины и даже длины, увеличение числа органелл, главным образом миофибрилл, а также нарастании числа ядер, что в конечном счете проявляется увеличением функциональной способности мышечного волокна. Радиоизотопным методом установлено, что увеличение числа ядер в мышечных волокнах в условиях гипертрофии достигается за счет деления клеток миосателлитов и последующего вхождения в миосимпласт дочерних клеток.
Увеличение числа миофибрилл осуществляется посредством синтеза актиновых и миозиновых белков свободными рибосомами и последующей сборки этих белков в актиновые и миозиновые миофиламенты параллельно с соответствующими филаментами саркомеров. В результате этого вначале происходит утолщение миофибрилл, а затем их расщепление и образование дочерних миофибрилл. Кроме того возможно образование новых актиновых и миозиновых миофиламентов не параллельно, а встык предшествующим миофибриллам, чем достигается их удлинение. Саркоплазматическая сеть и Т-канальцы в гипертрофирующемся волокне образуются за счет разрастания предшествующих элементов. При определенных видах мышечной тренировки может формироваться преимущественно красный тип мышечных волокон (у стайеров) или белый тип мышечных волокон (у спринтеров). Возрастная гипертрофия мышечных волокон интенсивно проявляется с началом двигательной активности организма (1-2 года), что обусловлено прежде всего усилением нервной стимуляции. В старческом возрасте, а также в условиях малой мышечной нагрузки наступает атрофия специальных и общих органелл, истончение мышечных волокон и снижение их функциональной способности.
Репаративная регенерация развивается после повреждения мышечных волокон. При этом способ регенерации зависит от величины дефекта. При значительных повреждениях на протяжении мышечного волокна миосателлиты в области повреждения и в прилежащих участках растормаживаются, усиленно пролиферируют, а затем мигрируют в область дефекта мышечного волокна, где выстраиваются в цепочки, формируя миотрубку. Последующая дифференцировка миотрубки приводит к восполнению дефекта и восстановлению целостности мышечного волокна. В условиях небольшого дефекта мышечного волокна на его концах, за счет регенерации внутриклеточных органелл, образуются мышечные почки, которые растут навстречу друг другу, а затем сливаются, приводя к закрытию дефекта. Однако, репаративная регенерация и восстановление целостности мышечных волокон могут осуществляться при определенных условиях: во-первых, при сохраненной двигательной иннервации мышечных волокон, во-вторых, если в область повреждения не попадают элементы соединительной ткани (фибробласты). Иначе на месте дефекта мышечного волокна развивается соединительно-тканный рубец.
Советским ученым А.Н. Студитским доказана возможность аутотрансплантации скелетной мышечной ткани и даже целых мышц при соблюдении определенных условий:
· механическое измельчение мышечной ткани трансплантата, с целью растормаживания клеток-сателлитов и последующей их пролиферации;
· помещение измельченной ткани в фасциальное ложе;
· подшивание двигательного нервного волокна к измельченному трансплантату;
· наличие сократительных движений мышц-антагонистов и синергистов.
Анатомически у новорожденных имеются все скелетные мышцы, но относительно веса тела они составляют всего 23% (у взрослого 44%). Количество мышечных волокон в мышцах такое же как у взрослого. Однако микроструктура мышечных волокон отличается: волокна меньше диаметром, в них больше ядер. По мере роста происходит утолщение и удлинение волокон. Это происходит за счет утолщения миофибрилл, оттесняющих ядра на периферию. Размеры мышечных волокон стабилизируются к 20 годам.
Мышцы у детей эластичнее, чем у взрослых. Т.е. быстрее укорачиваются при сокращении и удлиняются при расслаблении. Возбудимость и лабильность мышц новорожденных, ниже, чем взрослых, но с возрастом растет. У новорожденных даже во сне мышцы находятся в состоянии тонуса. Развитие различных групп мышц происходит неравномерно. В 4-5 лет более развиты мышцы предплечья, отстают в развитии мышцы кисти. Ускоренное согревание мышц кисти происходит в 6-7 лет. Причем разгибатели развиваются медленнее сгибателей. С возрастом изменяется соотношение тонуса мышц. В раннем детстве повышен тонус мышц кисти, разгибателей бедра т.д. постепенно распределение тонуса нормализуется.
Для сердца как органа характерна способность к регенерации путем регенераторной гипертрофии, при которой масса органа восстанавливается, но форма остается нарушенной. Подобное явление наблюдается после перенесенного инфаркта миокарда, когда масса сердца может восстановиться как целое, при этом на месте повреждения образуется соединительнотканный рубец, но орган гипертрофируется, т.е. нарушается форма. Происходит не только увеличение размеров кардиомиоцитов, но и пролиферация в основном в предсердиях и ушках сердца.
Ранее полагали, что дифференцировка кардиомиоцитов является необратимым процессом, связанным с полной потерей этими клетками способности к делению. Но на современном уровне многочисленные данные показывают то, что дифференцированные кардиомиоциты способны к синтезу ДНК и митозу. В исследовательских работах П.П. Румянцева и его учеников показано, что после экспериментального инфаркта миокарда левого желудочка сердца в клеточный цикл возвращается 60-70% предсердных кардиомиоцитов, возрастает число полиплоидных клеток, но это не компенсирует повреждение миокарда.
Установлено, что кардиомиоциты способны к митотическому делению (в том числе и клетки проводящей системы). В миокарде сердца особенно много одноядерных полиплоидных клеток с 16-32-кратным содержанием ДНК, но встречаются и двуядерные кардиомиоциты (13-14%) в основном октоплоидные.
В процессе регенерации сердечной мышечной ткани кардиомиоциты участвуют в процессе гиперплазии и гипертрофии, возрастает их плоидность, но уровень пролиферации клеток соединительной ткани в области повреждения оказывается в 20-40 раз выше. В фибробластах активизируется синтез коллагена, в результате чего репарация происходит путем рубцевания дефекта. Биологическое представление подобной адаптационной реакции соединительной ткани объясняется жизненной важностью сердечного органа, так как задержка с закрытием дефекта может привести к гибели.
Считалось, что у новорожденных, а возможно, и в раннем детском возрасте, когда способные к делению кардиомиоциты еще сохраняются, регенераторные процессы сопровождаются увеличением количества кардиомиоцитов. При этом у взрослых физиологическая регенерация осуществляется в миокарде в основном путем внутриклеточной регенерации, без увеличения количества клеток, т.е. в миокарде взрослого человека отсутствует пролиферация кардиомиоцитов. Но недавно получены данные о том, что в здоровом сердце человека 14 миоцитов из миллиона находится в состоянии митоза, завершающегося цитотомией, т.е. количество клеток не значительно, но увеличивается.
Применение современных методов клеточной биологии в клинических и экспериментальных исследованиях позволило перейти к выяснению клеточных и молекулярных механизмов повреждения и регенерации миокарда. Особенно интересны данные о том, что в перинекротических областях и в функционально перегруженном сердце происходит синтез эмбриональных миоакрдиальных белков и пептидов, а также белков, синтезирующихся во время клеточного цикла. Это подтверждает положение о сходстве механизмов регенерации и нормального онтогенеза.
Выяснилось также и то, что дифференцированные кардиомиоциты в культуре способны к активному митотическому делению, что, возможно, объясняется не полной утратой, а подавлением способности кардиомиоцитов возвращаться в клеточный цикл.
Важной задачей теоретической и практической кардиологии является разработка способов стимуляции восстановления поврежденного миокарда, т.е. индукции миокардиальной регенерации и уменьшения соединительнотканного рубца. Одно из направлений исследований предоставляет возможность переноса регуляторных генов, которые превращают фибробласты рубца в миобласты или трансфекция в кардиомиоциты генов, контролирующих рост новых клеток. Другим направлением является перенос в область повреждения фетальных скелетных и миокардиальных клеток, которые могли бы участвовать в восстановлении сердечной мышцы. Также проводятся эксперименты по трансплантации скелетной мышцы в сердце, показывающие образование в миокарде участков сокращающейся ткани и улучшающие функциональные показатели миокарда. Перспективным может быть лечение с применением факторов роста, оказывающих как прямое, так и косвенное воздействие на поврежденный миокард, например, улучшение ангиогенеза.
Гладкие мышечные ткани
По происхождению различают три группы гладких (или неисчерченных) мышечных тканей -- мезенхимные, эпидермальные и нейральные.
Мышечная ткань мезенхимного происхождения
Гистогенез. Стволовые клетки и клетки-предшественники гладкой мышечной ткани, будучи уже детерминированными, мигрируют к местам закладки органов. Дифференцируясь, они синтезируют компоненты матрикса и коллаген базальной мембраны, а также эластин. У дефинитивных клеток (миоцитов) синтетическая способность снижена, но не исчезает полностью.
Структурно-функциональной единицей гладкой, или неисчерченной, мышечной ткани является гладкомышечная клетка, или гладкий миоцит -- это веретеновидная клетка длиной 20--500 мкм, шириной 5--8 мкм. Ядро клетки палочковидное, находится в ее центральной части. Когда миоцит сокращается, его ядро изгибается и даже закручивается. Органеллы общего значения, среди которых много митохондрий, сосредоточены в цитоплазме около полюсов ядра. Аппарат Гольджи и гранулярная эндо плазматическая сеть развиты слабо, что свидетельствует о малой активности синтетических функций. Рибосомы в большинстве своем расположены свободно.
Филаменты актина образуют в цитоплазме трехмерную сеть, вытянутую преимущественно продольно, точнее косо-продольно. Концы филаментов скреплены между собой и с плазмолеммой специальными сшивающими белками. Эти участки хорошо видны на электронных микрофотографиях как плотные тельца.
Миозиновые филаменты находятся в деполимеризованном состоянии. Мономеры миозина располагаются рядом с филаментами актина. Сигнал к сокращению обычно поступает по нервным волокнам. Медиатор, который выделяется из их терминалей, изменяет состояние плазмолеммы. Она образует впячивания -- кавеолы, в которых концентрируются ионы кальция. Кавеолы отшнуровываются в сторону цитоплазмы в виде пузырьков (здесь из пузырьков освобождается кальций). Это влечет за собой как полимеризацию миозина, так и взаимодействие миозина с актином. Актиновые филаменты смещаются друг другу навстречу, плотные пятна сближаются, усилие передается на плазмолемму, и вся клетка укорачивается. Когда поступление сигналов со стороны нервной системы прекращается, ионы кальция эвакуируются из кавеол, миозин деполимеризуется и «миофибриллы» распадаются. Таким образом, актино-миозиновые комплексы существуют в гладких миоцитах только в период сокращения.
Гладкие миоциты располагаются без заметных межклеточных пространств и разделены базальной мембраной. На отдельных участках в ней образуются «окна», поэтому плазмолеммы соседних миоцитов сближаются. Здесь формируются нексусы, и между клетками возникают не только механические, но и метаболические связи. Поверх «чехликов» из базальной мембраны между миоцитами проходят эластические и ретикулярные волокна, объединяющие клетки в единый тканевой комплекс. Ретикулярные волокна проникают в щели на концах миоцитов, закрепляются там и передают усилие сокращения клетки всему их объединению.
Регенерация. Физиологическая регенерация гладкой мышечной ткани проявляется в условиях повышенных функциональных нагрузок. Наиболее отчетливо это видно в мышечной оболочке матки при беременности. Такая регенерация осуществляется не столько на тканевом, сколько на клеточном уровне: миоциты растут, в цитоплазме активизируются синтетические процессы, количество миофиламентов увеличивается (рабочая гипертрофия клеток). Не исключена, однако, и пролиферация клеток (т.е. гиперплазия).
В составе органов миоциты объединяются в пучки, между которыми располагаются тонкие прослойки соединительной ткани. В эти прослойки вплетаются ретикулярные и эластические волокна, окружающие миоциты. В прослойках проходят кровеносные сосуды и нервные волокна. Терминали последних оканчиваются не непосредственно на миоцитах, а между ними. Поэтому после поступления нервного импульса медиатор распространяется диффузно, возбуждая сразу многие клетки. Гладкая мышечная ткань мезенхимного происхождения представлена главным образом в стенках кровеносных сосудов и многих трубчатых внутренних органов, а также образует отдельные мелкие мышцы.
Гладкая мышечная ткань в составе конкретных органов имеет неодинаковые функциональные свойства. Это обусловлено тем, что на поверхности органов имеются разные рецепторы к конкретным биологически активным веществам. Поэтому и на многие лекарственные препараты их реакция неодинакова.
Гладкая мышечная ткань эпидермального происхождения
Миоэпителиальные клетки развиваются из эпидермального зачатка. Они встречаются в потовых, молочных, слюнных и слезных железах и имеют общих предшественников с железистыми секреторными клетками. Миоэпителиальные клетки непосредственно прилежат к собственно эпителиальным и имеют общую с ними базальную мембрану. При регенерации те и другие клетки восстанавливаются из общих малодифференцированных предшественников. Большинство миоэпителиальных клеток имеют звездчатую форму. Эти клетки нередко называют корзинчатыми: их отростки охватывают концевые отделы и мелкие протоки желез. В теле клетки располагаются ядро и органеллы общего значения, а в отростках -- сократительный аппарат, организованный, как и в клетках мышечной ткани мезенхимного типа.
Гладкая мышечная ткань нейрального происхождения
Миоциты этой ткани развиваются из клеток нейрального зачатка в составе внутренней стенки глазного бокала. Тела этих клеток располагаются в эпителии задней поверхности радужки. Каждая из них имеет отросток, который направляется в толщу радужки и ложится параллельно ее поверхности. В отростке находится сократительный аппарат, организованный так же, как и во всех гладких миоцитах. В зависимости от направления отростков (перпендикулярно или параллельно краю зрачка) миоциты образуют две мышцы -- суживающую и расширяющую зрачок.
Заключение
Как уже было отмечено, мышечные ткани - это группа тканей организма различного происхождения, объединяемых по признаку сократимости: поперечнополосатая (скелетная и сердечная), гладкая, а также специализированные сократимые ткани - эпителиально-мышечная и нейроглиальная, входящая в состав радужки глаза.
Поперечнополосатая скелетная мышечная ткань возникает из миотомов, входящих в состав элементов сегментированной мезодермы - сомитов.
Гладкая мышечная ткань человека и позвоночных животных развивается в составе производных мезенхимы, так же как и ткани внутренней среды. Однако для всех мышечных тканей характерно сходное обособление в составе эмбрионального зачатка в виде клеток веретенообразной формы - мышцеобразовательных клеток, или миобластов.
Сокращение мышечного волокна заключается в укорочении миофибрилл в пределах каждого саркомера. Толстые (миозиновые) и тонкие (актиновые) нити, в расслабленном состоянии связанные только концевыми отделами, в момент сокращения осуществляют скользящие движения навстречу друг другу. Выделение необходимой для сокращения энергии происходит в результате превращения АТФ в АДФ под влиянием миозина. Ферментная активность миозина проявляется при условии оптимального содержания Са2+, которые накапливаются в саркоплазматической сети.
Список литературы
1. Гистология. Под редакцией Ю.И. Афанасьевой, Н.А. Юриной. М.: “Медицина”, 1999 г.
2. Р. Эккерт, Д. Рендел, Дж. Огастин “Физиология животных” - 1 т. М.: “Мир”, 1981 г.
3. К.П. Рябов “Гистология с основами эмбриологии” Минск: “Высшая школа”, 1990 г.
4. Гистология. Под редакцией Улумбекова, проф. Ю.А. Челышева. М.: 1998 г.
5. Гистология. Под редакцией В.Г. Елисеева. М.: “Медицина”, 1983 г.
Размещено на Allbest.ru
...Подобные документы
Структурные особенности мышечных тканей. Изучение механизма мышечного сокращения и аппарата передачи возбуждения. Гистогенез и регенерация мышечной ткани. Принципы работы сократительных, проводящих и секреторных кардиомиоцитов сердечной мышечной ткани.
шпаргалка , добавлен 14.11.2010
Изучение видов и функций различных тканей человека. Задачи науки гистологии, которая изучает строение тканей живых организмов. Особенности строения эпителиальной, нервной, мышечной ткани и тканей внутренней среды (соединительной, скелетной и жидкой).
презентация , добавлен 08.11.2013
Изучение особенностей строения тканей животных, функционирование и разновидности. Проведение исследования характерной черты строения соединительной и нервной тканей. Структура плоской, кубической, мерцательной и железистой эпителии. Виды мышечной ткани.
презентация , добавлен 08.02.2015
Общая характеристика и возрастные особенности хрящевой ткани. Виды хрящевой и костной ткани. Общая характеристика и возрастные особенности костной ткани. Особенности строения мышечной ткани в детском и в пожилом возрасте. Скелетная мышечная ткань.
презентация , добавлен 07.02.2016
Класификация тканей, виды эпителиальных тканей, их строение и функции. Опорная, трофическая и защитная функция соединительных тканей. Функции нервной и мышечной тканей. Понятие об органах и системах органов, их индивидуальные, половые, возрастные отличия.
реферат , добавлен 11.09.2009
Эпителиальная ткань, ее регенерационная способность. Соединительные ткани, участвующие в поддержании гомеостаза внутренней среды. Клетки кровы и лимфы. Поперечнополосатые и сердечные мышечные ткани. Функции нервных клеток и тканей животных организмов.
реферат , добавлен 16.01.2015
Изучение видов тканей животных, а также функций, которые они выполняют. Особенности строения эпителиальной, соединительной, мышечной и нервной группы тканей. Определение месторасположения каждой группы и значения для жизнедеятельности организма животного.
презентация , добавлен 18.10.2013
Общее понятие и разновидности колебаний. Характеристика процессов растяжения (сжатия), сдвига, изгиба, кручения. Механические свойства костной и сосудистой тканей. Специфика мышечной ткани, основные режимы работы мышц – изометрический и изотонический.
контрольная работа , добавлен 19.03.2014
Клетка как основная структурная единица организма. Описание ее строения, жизненных и химических свойств. Строение и функции эпителиальной и соединительной, мышечной и нервной тканей. Органы и перечень системы органов человека, их назначение и функции.
презентация , добавлен 19.04.2012
Физиология и биохимия мышечной деятельности как важная составляющая обмена веществ в организме. Типы мышечной ткани и соответственно мышц, различающихся по структуре мышечных волокон, характеру иннервации. Влияние физических нагрузок разной интенсивности.
Организм всех животных, в том числе и человека, состоит из четырех типов тканей: эпителиальной, нервной, соединительной и мышечной. О последней и пойдет речь в данной статье.
Разновидности мышечной ткани
Она бывает трех видов:
- поперечно-полосатая;
- гладкая;
- сердечная.
Функции мышечных тканей разных видов несколько отличаются. Да и строение тоже.
Где находятся мышечные ткани в организме человека?
Мышечные ткани разных видов занимают различное местоположение в организме животных и человека. Так, из сердечной мускулатуры, как понятно из названия, построено сердце.
Из поперечно-полосатой мышечной ткани образуются скелетные мускулы.
Гладкие мышцы выстилают изнутри полости органов, которым необходимо сокращаться. Это, к примеру, кишечник, мочевой пузырь, матка, желудок и т.д.
Структура мышечной ткани разных видов различается. О ней поговорим подробнее дальше.
Как устроена мышечная ткань?
Она состоит из больших по размеру клеток — миоцитов. Они также еще называются волокнами. Клетки мышечной ткани обладают несколькими ядрами и большим количеством митохондрий — органоидов, отвечающих за выработку энергии.
Кроме того, строение мышечной ткани человека и животных предусматривает наличие небольшого количества межклеточного вещества, содержащего коллаген, который придает мышцам эластичность.
Давайте рассмотрим строение и функции мышечных тканей разных видов по отдельности.
Структура и роль гладкой мышечной ткани
Данная ткань контролируется вегетативной нервной системой. Поэтому человек не может сокращать сознательно мышцы, построенные из гладкой ткани.
Формируется она из мезенхимы. Это разновидность эмбриональной соединительной ткани.
Сокращается данная ткань намного менее активно и быстро, чем поперечно-полосатая.
Гладкая ткань построена из миоцитов веретеновидной формы с заостренными концами. Длина данных клеток может составлять от 100 до 500 микрометров, а толщина — около 10 микрометров. Клетки данной ткани являются одноядерными. Ядро расположено в центре миоцита. Кроме того, хорошо развиты такие органоиды, как агранулярная ЭПС и митохондрии. Также в клетках гладкой мышечной ткани присутствует большое количество включений из гликогена, которые представляют собой запасы питательных веществ.
Элементом, который обеспечивает сокращение мышечной ткани данного вида, являются миофиламенты. Они могут быть построены из двух сократительных белков: актина и миозина. Диаметр миофиламентов, которые состоят из миозина, составляет 17 нанометров, а тех, которые построены из актина — 7 нанометров. Существуют также промежуточные миофиламенты, диаметр которых составляет 10 нанометров. Ориентация миофибрилл продольная.
В состав мышечной ткани данного вида также входит межклеточное вещество из коллагена, которое обеспечивает связь между отдельными миоцитами.
Функции мышечных тканей этого вида:
- Сфинктерная. Заключается в том, что из гладких тканей устроены круговые мышцы, регулирующие переход содержимого из одного органа в другой или из одной части органа в другую.
- Эвакуаторная. Заключается в том, что гладкие мышцы помогают организму выводить ненужные вещества, а также принимают участие в процессе родов.
- Создание просвета сосудов.
- Формирование связочного аппарата. Благодаря ему многие органы, такие как, например, почки, удерживаются на своем месте.
Теперь давайте рассмотрим следующий вид мышечной ткани.
Поперечно-полосатая
Она регулируется соматической нервной системой. Поэтому человек может сознательно регулировать работу мышц данного вида. Из поперечно-полосатой ткани формируется скелетная мускулатура.
Данная ткань состоит из волокон. Это клетки, которые обладают множеством ядер, расположенных ближе к плазматической мембране. Кроме того, в них находится большое количество гликогеновых включений. Хорошо развиты такие органоиды, как митохондрии. Они находятся вблизи сократительных элементов клетки. Все остальные органеллы локализуются неподалеку от ядер и развиты слабо.
Структурами, благодаря которым поперечно-полосатая ткань сокращается, являются миофибриллы. Их диаметр составляет от одного до двух микрометров. Миофибриллы занимают большую часть клетки и расположены в ее центре. Ориентация миофибрилл продольная. Они состоят из светлых и темных дисков, которые чередуются, что и создает поперечную "полосатость" ткани.
Функции мышечных тканей данного вида:
- Обеспечивают перемещение тела в пространстве.
- Отвечают за передвижение частей тела друг относительно друга.
- Способны к поддержанию позы организма.
- Участвуют в процессе регуляции температуры: чем активнее сокращаются мышцы, тем выше температура. При замерзании поперечно-полосатые мышцы могут начать сокращаться непроизвольно. Этим и объясняется дрожь в теле.
- Выполняют защитную функцию. Особенно это касается мышц брюшного пресса, которые защищают многие внутренние органы от механических повреждений.
- Выступают в роли депо воды и солей.
Сердечная мышечная ткань
Данная ткань похожа одновременно и на поперечно-полосатую, и на гладкую. Как и гладкая, она регулируется вегетативной нервной системой. Однако сокращается она так же активно, как и поперечно-полосатая.
Состоит она из клеток, называющихся кардиомиоцитами.
Функции мышечной ткани данного вида:
- Она всего одна: обеспечение передвижения крови по организму.
У позвоночных животных и человека различают три разных по строению группы мышц :
- поперечно-полосатые мышцы скелета;
- поперечно-полосатая мышца сердца;
- гладкие мышцы внутренних органов, сосудов и кожи.
Рис. 1. Виды мышц человека
Гладкие мышцы
Из двух видов мышечной ткани (поперечно-полосатой и гладкой) гладкая мышечная ткань находится на более низкой ступени развития и присуща низшим животным.
Образуют мышечный слой стенок желудка, кишечника, мочеточников, бронхов, кровеносных сосудов и других полых органов. Они состоят из веретенообразных мышечных волокон и не имеют поперечной исчерченности, так как миофибриллы в них расположены менее упорядоченно. В гладких мышцах отдельные клетки соединяются между собой специальными участками наружных мембран - нексусами . За счет этих контактов потенциалы действия распространяются с одного мышечного волокна на другое. Поэтому в реакцию возбуждения быстро вовлекается вся мышца.
Гладкие мышцы осуществляют движения внутренних органов, кровеносных и лимфатических сосудов. В стенках внутренних органов они, как правило, располагаются в виде двух слоев: внутреннего кольцевого и наружного продольного. В стенках артерии они формируют спиралевидные структуры.
Характерной особенностью гладких мышц является их способность к спонтанной автоматической деятельности (мышцы желудка, кишечника, желчного пузыря, мочеточников). Это свойство регулируется нервными окончаниями. Гладкие мышцы пластичны, т.е. способны сохранять приданную растяжением длину без изменения напряжения. Скелетная мышца, наоборот, обладает малой пластичностью и эту разницу легко установить в следующем опыте: если растянуть с помощью грузов и гладкую и поперечно-полосатую мышцы и снять груз, то скелетная мышца сразу же после этого укорачивается до первоначальной длины, а гладкая мышца долгое время может находиться в растянутом состоянии.
Такое свойство гладких мышц имеет большое значение для функционирования внутренних органов. Именно пластичность гладких мышц обеспечивает лишь небольшое изменение давления внутри мочевого пузыря при его наполнении.

Рис. 2. А. Волокно скелетной мышцы, клетка сердечной мышцы, гладкая мышечная клетка. Б. Саркомер скелетной мышцы. В. Строение гладкой мышцы. Г. Механограмма скелетной мышцы и мышцы сердца.
Гладким мышцам присущи те же основные свойства, что и поперечнополосатым скелетным мышцам, но и некоторые особые свойства:
- автоматия, т.е. способность сокращаться и расслабляться без внешних раздражений, а за счет возбуждений, возникающих в них самих;
- высокая чувствительность к химическим раздражителям;
- выраженная пластичность;
- сокращение в ответ на быстрое растяжение.
Сокращение и расслабление гладких мышц происходит медленно. Это способствует наступлению перестальтических и маятникообразных движений органов пищеварительного тракта, что приводит к перемещению пищевого комка. Длительное сокращение гладких мышц необходимо в сфинктерах полых органов и препятствует выходу содержимого: желчи в желчном пузыре, мочи в мочевом пузыре. Сокращение гладкомышечных волокон совершается независимо от нашего желания, под воздействием внутренних, не подчиненных сознанию причин.
Поперечно-полосатые мышцы
Поперечно-полосатые мышцы располагаются на костях скелета и сокращением приводят в движение отдельные суставы и все тело. образуют тело, или сому, поэтому их еще называют соматическими, а иннервирующую их систему — соматической нервной системой.
Благодаря деятельности скелетной мускулатуры осуществляется передвижение тела в пространстве, разнообразная работа конечностей, расширение грудной клетки при дыхании, движение головы и позвоночника, жевание, мимика лица. Насчитывается более 400 мышц. Общая масса мышц составляет 40% веса. Обычно средняя часть мышцы состоит из мышечной ткани и образует брюшко. Концы мышц — сухожилия построены из плотной соединительной ткани; они соединяются с костями при помощи надкостницы, но могут прикрепляться и к другой мышце, и к соединительному слою кожи. В мышце мышечные и сухожильные волокна объединяются в пучки при помощи рыхлой соединительной ткани. Между пучками располагаются нервы и кровеносные сосуды. пропорциональна количеству волокон, составляющих брюшко мышцы.

Рис. 3. Функции мышечной ткани
Некоторые мышцы проходят только через один сустав и при сокращении приводят его в движение — односуставные мышцы. Другие мышцы проходят через два или несколько суставов — многосуставные, они производят движение в нескольких суставах.
При концы мышцы, прикрепленные к костям, приближаются друг к другу, а размеры мышцы (длина) уменьшается. Кости, соединенные суставами, действуют как рычаги.
Изменяя положение костных рычагов, мышцы действуют на суставы. При этом каждая мышца влияет на сустав только в одном направлении. У одноосного сустава (цилиндрический, блоковидный) имеются две действующие на него мышцы или группы мышц, являющиеся антагонистами: одна мышца — сгибатель, другая — разгибатель. В то же время на каждый сустав в одном направлении действует, как правило, две мышцы и более, являющиеся синергистами (синергизм — совместное действие).
У двуосного сустава (эллипсоидный, мышелковый, седловидный) мышцы группируются соответственно двум его осям, вокруг которых совершаются движения. К шаровидному суставу, имеющему три оси движения (многоосный сустав), мышцы прилежат со всех сторон. Так, например, в плечевом суставе имеются мышцы-сгибатели и разгибатели (движения вокруг фронтальной оси), отводящие и приводящие (сагиттальная ось) и вращатели вокруг продольной оси, кнутри и кнаружи. Различают три вида работы мышц: преодолевающую, уступающую и удерживающую.
Если благодаря сокращению мышцы меняется положение части тела, то преодолевается сила сопротивления, т.е. выполняется преодолевающая работа. Работа, при которой сила мышцы уступает действию силы тяжести и удерживаемого груза, называется уступающей. В этом случае мышца функционирует, однако она не укорачивается, а удлиняется, например, когда невозможно поднять или удержать на весу тело, имеющее большую массу. При большом усилии мышц приходится опустить это тело на какую-нибудь поверхность.
Удерживающая работа выполняется благодаря сокращению мышц, тело или груз удерживается в определенном положении без перемещения в пространстве, например человек держит груз, не двигаясь. При этом мышцы сокращаются без изменения длины. Сила сокращения мышц уравновешивает массу тела и груза.
Когда мышца, сокращаясь, перемешает тело или его части в пространстве, они выполняют преодолевающую или уступающую работу, которая является динамической. Статистической является удерживающая работа, при которой не происходит движений всего тела или его части. Режим, при котором мышца может свободно укорачиваться, называется изотоническим (не происходит изменения напряжения мышцы и меняется только ее длина). Режим, при котором мышца не может укоротиться, называется изометрическим — меняется только напряжение мышечных волокон.

Рис. 4. Мышцы человека
Строение поперечно-полосатых мышц
Скелетные мышцы состоят из большого числа мышечных волокон, которые объединяются в мышечные пучки.
В одном пучке содержится 20-60 волокон. Мышечные волокна представляют собой клетки цилиндрической формы длиной 10-12 см и диаметром 10-100 мкм.
Каждое мышечное волокно имеет оболочку (сарколемму) и цитоплазму (саркоплазму). В саркоплазме находятся все компоненты животной клетки и вдоль оси мышечного волокна располагаются тонкие нити - миофибриллы, Каждая миофибрилла состоит из протофибрилл, в состав которых вкючены нити белков миозина и актина, являющихся сократительным аппаратом мышечного волокна. Миофибриллы разделены между собой перегородками, которые называются Z-мембранами, на участки - саркомеры. На обоих концах саркомеров к Z-мембране прикреплены тонкие актиновые нити, а в середине расположены толстые миозиновые нити. Нити актина своими концами частично входят между миозиновыми нитями. В световом микроскопе нити миозина выглядят в виде светлой полоски в темном диске. При электронной микроскопии скелетные мышцы выглядят исчерченными (поперечно-полосатыми).

Рис. 5. Поперечные мостики: Ак — актин; Мз — миозин; Гл — головка; Ш — шейка
На боковых сторонах миозиновой нити имеются выступы, получившие название поперечных мостиков (рис. 5), которые расположены под углом 120° по отношению к оси миозиновой нити. Актиновые филаменты выглядят в виде двойной нити, закрученной в двойную спираль. В продольных бороздках актиновой спирали находятся нити белка тропомиозина, к которым присоединен белок тропонин. В состоянии покоя молекулы белка тропомиозина расположены таким образом, чтобы предотвращать прикрепление поперечных мостиков миозина к актиновым нитям.

Рис. 6. А — организация цилиндрических волокон в скелетной мышце, прикрепленной к костям сухожилиями. Б — структурная организация филаментов в волокне скелетной мышцы, создающая картину поперечных полос.

Рис. 7. Строение актина и миозина
Во многих местах поверхностная мембрана углубляется в виде микротрубок внутрь волокна, перпендикулярно его продольной оси, образуя систему поперечных трубочек (Т-система). Параллельно миофибриллам и перпендикулярно поперечным трубочкам между миофибрилл расположена система продольных трубочек (саркоплазматический ретикулум). Концевые расширения этих трубочек - терминальные цистерны - подходят очень близко к поперечным трубочкам, образуя совместно с ними так называемые триады. В цистернах сосредоточено основное количество внутриклеточного кальция.
Механизм сокращения скелетной мышцы
Мышечная ткань состоит из клеток, называемых мышечными волокнами. Снаружи волокно окружено оболочкой — сарколеммой. Внутри сарколеммы содержится цитоплазма (саркоплазма), содержащая ядра и митохондрии. В ней содержится огромное количество сократительных элементов, называемых миофибриллами. Миофибриллы проходят от одного конца мышечного волокна до другого. Они существуют сравнительно короткий срок — около 30 суток, после чего и происходит их полная смена. В мышцах идет интенсивный синтез белка, необходимый для образования новых миофибрилл.
Мышечное волокно содержит большое количество ядер, которые располагаются непосредственно под сарколеммой, поскольку основная часть мышечного волокна занята миофибриллами. Именно наличие большого числа ядер обеспечивает синтез новых миофибрилл. Такая быстрая смена миофибрилл обеспечивает высокую надежность физиологических функций мышечной ткани.

Рис. 7. А — схема организации саркоплазматического ретикулума, поперечных трубочек и миофибрилл. Б — схема анатомической структуры поперечных трубочек и саркоплазматического ретикулума в индивидуальном волокне скелетной мышцы. В — роль саркоплазматического ретикулума в механизме сокращения скелетной мышцы
Каждая миофибрилла состоит из правильно чередующихся светлых и темных участков. Эти участки, обладая разными оптическими свойствами, создают поперечную исчерченность мышечной ткани.
В скелетной мышце сокращение вызывается поступлением к ней импульса по нерву. Передача нервного импульса с нерва на мышцу осуществляется через нервно-мышечный синапс (контакт).
Одиночный нервный импульс, или однократное раздражение, приводит к элементарному сократительному акту — одиночному сокращению. Начало сокращения не совпадает с моментом нанесения раздражения, поскольку существует скрытый, или латентный, период (интервал между нанесением раздражения и началом сокращения мышцы). В этот период происходит развитие потенциала действия, активация ферментных процессов и распад АТФ. После этого начинается сокращение. Распад АТФ в мышце приводит к превращению химической энергии в механическую. Энергетические процессы всегда сопровождаются выделением тепла и тепловая энергия обычно является промежуточной между химической и механическими энергиями. В мышце же химическая энергия превращается непосредственно в механическую. Но тепло в мышце образуется и за счет укорочения мышцы, и во время ее расслабления. Тепло, образующееся в мышцах, играет большую роль в поддержании температуры тела.
В отличие от сердечной мышцы, которая обладает свойством автоматики, т.е. она способна сокращаться под влиянием импульсов, возникающих в ней самой, и в отличие от гладкой мускулатуры, также способной к сокращению без поступления сигналов извне, скелетная мышца сокращается только при поступлении к ней сигналов из . Непосредственно сигналы к мышечным волокнам поступают по аксонам двигательных клеток, расположенным в передних рогах серого вещества спинного мозга (мотонейронам).
Рефлекторный характер деятельности мышц и координация мышечных сокращений
Скелетные мышцы в отличие от гладких способны совершать произвольные быстрые сокращения и производить этим значительную работу. Рабочим элементом мышцы является мышечное волокно. Типичное мышечное волокно представляет собой структуры с несколькими ядрами, отодвинутыми на периферию массой сократительных миофибрилл.
Мышечные волокна обладают тремя основными свойствами:
- возбудимостью — способностью отвечать на действия раздражителя генерацией потенциала действия;
- проводимостью — способностью проводить волну возбуждения вдоль всего волокна в обе стороны от точки раздражения;
- сократимостью — способностью сокращаться или изменять напряжение при возбуждении.
В физиологии имеется понятие двигательной единицы, под которой подразумевается один двигательный нейрон и все мышечные волокна, которые этот нейрон иннервирует. Двигательные единицы бывают разными по объему: от 10 мышечных волокон на единицу для мышц, выполняющих точные движения, до 1000 и более волокон на двигательную единицу для мышц «силовой направленности». Характер работы скелетных мышц может быть различным: статическая работа (поддержание позы, удержание груза) и динамическая работа (перемещение тела или груза в пространстве). Мышцы участвуют также в передвижении крови и лимфы в организме, выработке тепла, актах вдоха и выдоха, являются своеобразными депо для воды и солей, защищают внутренние органы, например мышцы брюшной стенки.
Для скелетной мышцы характерны два основных режима сокращения — изометрический и изотонический.
Изометрический режим проявляется в том, что в мышце во время ее активности нарастает напряжение (генерируется сила), но из-за того, что оба конца мышцы фиксированы (например, при попытке поднять очень большой груз), — она не укорачивается.
Изотонический режим проявляется в том, что мышца первоначально развивает напряжение (силу), способное поднять данный груз, а потом мышца укорачивается — меняет свою длину, сохраняя напряжение, равное весу удерживаемого груза. Чисто изометрического или изотонического сокращения практически наблюдать нельзя, но существуют приемы так называемой изометрической гимнастики, когда спортсмен напрягает мышцы без изменения длины. Эти упражнения в большей мере развивают силу мышц, чем упражнения с изотоническими элементами.
Сократительный аппарат скелетной мышцы представлен миофибриллами. Каждая миофибрилла диаметром 1 мкм состоит из нескольких тысяч протофибрилл — тонких, удлиненных полимеризированных молекул белков миозина и актина. Миозиновые нити в два раза тоньше актиновых, и в состоянии покоя мышечного волокна актиновые нити свободными кольцами входят между миозиновыми нитями.
В передаче возбуждения большую роль играют ионы кальция, которые входят в межфибриллярное пространство и запускают механизм сокращения: взаимное втягивание относительно друг друга актиновых и миозиновых нитей. Втягивание нитей происходит при обязательном участии АТФ. В активных центрах, расположенных на одном из концов миозиновых нитей, АТФ расщепляется. Энергия, выделяемая при расщеплении АТФ, преобразуется в движение. В скелетных мышцах запас АТФ невелик — всего на 10 одиночных сокращений. Поэтому необходим постоянный ре- синтез АТФ, который идет тремя путями: первый — за счет запасов креатинфосфата, которые ограничены; второй — гликолитический путь при анаэробном расщеплении глюкозы, когда на одну молекулу глюкозы образуется две молекулы АТФ, но одновременно образуется молочная кислота, которая тормозит активность гликолитических ферментов, и наконец третий — аэробное окисление глюкозы и жирных кислот в цикле Кребса, совершающееся в митохондриях и образующее 38 молекул АТФ на 1 молекулу глюкозы. Последний процесс наиболее экономичный, но очень медленный. Постоянная тренировка активизирует третий путь окисления, в результате чего повышается выносливость мышц к длительным нагрузкам.
Мышечные ткани представляют собой группу тканей различного происхождения и строения, объединенных на основании общего признака - выраженной сократительной способности, благодаря которой они могут выполнять свою основную функцию - перемещать тело или его части в пространстве.
Важнейшие свойства мышечных тканей. Структурные элементы мышечных тканей (клетки, волокна) обладают удлиненной формой и способны к сокращению благодаря мощному развитию сократительного аппарата. Для последнего характерно высокоупорядоченное расположение актиновых и миозиновых миофиламентов, создающее оптимальные условия для их взаимодействия. Это достигается связью сократимых структур с особыми элементами цитоскелета и плазмолеммой (сарколеммой), выполняющими опорную функцию. В части мышечных тканей миофиламенты образуют органеллы специального значения - миофибриллы. Для мышечного сокращения требуется значительное количество энергии, поэтому в структурных элементах мышечных тканей имеется большое количество митохондрий и трофических включений (липидных капель, гранул гликогена), содержащих субстраты - источники энергии. Поскольку мышечное сокращение протекает с участием ионов кальция, в мышечных клетках и волокнах хорошо развиты структуры, осуществляющие его накопление и выделение - агранулярная эндоплазматическая сеть (саркоплазматическая сеть), кавеолы.
Классификация мышечных тканей основана на признаках их (а) строения и функции (морфофункциональная классификация) и (б) происхождения (гистогенетическая классификация).
Морфофункциональная классификация мышечных тканей выделяет поперечнополосатые (исчерченные) мышечные ткани и гладкую мышечную ткань. Поперечнополосатые мышечные ткани образованы структурными элементами (клетками, волокнами), которые обладают поперечной исчерченностью вследствие особого упорядоченного взаиморасположения в них актиновых и миозиновых миофиламентов. К поперечнополосатым мышечным тканям относят скелетную и сердечную мышечную ткани. Гладкая мышечная ткань состоит из клеток, не обладающих поперечной исчерченностью. Наиболее распространенным видом этой ткани является гладкая мышечная ткань, входящая в состав стенки различных органов (бронхов, желудка, кишки, матки, маточной трубы, мочеточника, мочевого пузыря и сосудов).
Гистогенетическая классификация мышечных тканей выделяет три основных типа мышечных тканей: соматический (скелетная мышечная ткань), целомический (сердечная мышечная ткань) и мезенхимный (гладкая мышечная ткань внутренних органов), а также два дополнительных: миоэпителиальные клетки (видоизмененные эпителиальные сократимые клетки в концевых отделах и мелких выводных протоках некоторых желез) и мионейральные элементы (сократимые клетки нейрального происхождения в радужке глаза).
Скелетная поперечнополосатая (исчерченная) мышечная ткань по своей массе превышает любую другую ткань организма и является самой распространенной мышечной тканью тела человека. Она обеспечивает перемещение тела и его частей в пространстве и поддержание позы (входит в состав локомоторного аппарата), образует глазодвигательные мышцы, мышцы стенки полости рта, языка, глотки, гортани. Аналогичное строение имеет нескелетная висцеральная исчерченная мышечная ткань, которая обнаруживается в верхней трети пищевода, входит в состав наружных анального и уретрального сфинктеров.
Скелетная поперечнополосатая мышечная ткань развивается в эмбриональном периоде из миотомов сомитов, дающих начало активно делящимся миобластам - клеткам, которые располагаются цепочками и сливаются друг с другом в области концов с образованием мышечных трубочек (миотубул) , превращающихся в мышечные волокна. Такие структуры, образованные единой гигантской цитоплазмой и многочисленными ядрами, в отечественной литературе традиционно именуют симпластами (в данном случае - миосимпластами), однако этот термин отсутствует в принятой международной терминологии. Некоторые миобласты не сливаются с другими, располагаясь на поверхности волокон и давая начало миосателлитоцитам - мелким клеткам, которые являются камбиальными элементами скелетной мышечной ткани. Скелетная мышечная ткань образована собранными в пучки поперечнополосатыми мышечными волокнами (рис. 87), являющимися ее структурно-функциональными единицами.
Мышечные волокна скелетной мышечной ткани представляют собой цилиндрические образования вариабельной длины (от миллиметров до 10-30 см). Их диаметр также широко варьирует в зависимости от принадлежности к определенной мышце и типу, функционального состояния, степени функциональной нагрузки, состояния питания
и других факторов. В мышцах мышечные волокна образуют пучки, в которых они лежат параллельно и, деформируя друг друга, часто приобретают неправильную многогранную форму, что особенно хорошо видно на поперечных срезах (см. рис. 87). Между мышечными волокнами располагаются тонкие прослойки рыхлой волокнистой соединительной ткани, несущие сосуды и нервы - эндомизий. Поперечная исчерченность скелетных мышечных волокон обусловлена чередованием темных анизотропных дисков (полос А) и светлых изотропных дисков (полос I). Каждый изотропный диск рассекается надвое тонкой темной линией Z - телофрагмой (рис. 88). Ядра мышечного волокна - сравнительно светлые, с 1-2 ядрышками, диплоидные, овальные, уплощенные - лежат на его периферии под сарколеммой и располагаются вдоль волокна. Снаружи сарколемма покрыта толстой базальной мембраной, в которую вплетаются ретикулярные волокна.
Миосателлитоциты (клетки-миосателлиты) - мелкие уплощенные клетки, располагающиеся в неглубоких вдавлениях сарколеммы мышечного волокна и покрытые общей базальной мембраной (см. рис. 88). Ядро миосателлитоцита - плотное, относительно крупное, органеллы мелкие и немногочисленные. Эти клетки активируются при повреждении мышечных волокон и обеспечивают их репаративную регенерацию. Сливаясь с остальной частью волокна при усиленной нагрузке, миосателлитоциты участвуют в его гипертрофии.
Миофибриллы образуют сократительный аппарат мышечного волокна, располагаются в саркоплазме по ее длине, занимая центральную часть, и отчетливо выявляются на поперечных срезах волокон в виде мелких точек (см. рис. 87 и 88).
Миофибриллы обладают собственной поперечной исчерченностью, причем в мышечном волокне они располагаются столь упорядоченно, что изотропные и анизотропные диски разных миофибрилл совпадают между собой, обусловливая поперечную исчерченность всего волокна. Каждая миофибрилла образована тысячами повторяющихся последовательно связанных между собой структур - саркомеров.
Саркомер (миомер) является структурно-функциональной единицей миофибриллы и представляет собой ее участок, расположенный между двумя телофрагмами (линиями Z). Он включает анизотропный диск и две половины изотропных дисков - по одной половине с каждой стороны (рис. 89). Саркомер образован упорядоченной системой толстых (миозиновых) и тонких (актиновьх) миофиламентов. Толстые миофиламенты связаны с мезофрагмой (линией М) и сосредоточены в анизотропном диске,
а тонкие миофиламенты прикреплены к телофрагмам (линиям Z), образуют изотропные диски и частично проникают в анизотропный диск между толстыми нитями вплоть до светлой полосы Н в центре анизотропного диска.
Механизм мышечного сокращения описывается теорией скользящих нитей, согласно которой укорочение каждого саркомера (а, следовательно, миофибрилл и всего мышечного волокна) при сокращении происходит благодаря тому, что в результате взаимодействия актина и миозина в присутствии кальция и АТФ тонкие нити вдвигаются в промежутки между толстыми без изменения их длины. При этом ширина анизотропных дисков не меняется, а ширина изотропных дисков и полос Н - уменьшается. Строгая пространственная упорядоченность взаимодействия множества толстых и тонких миофиламентов в саркомере определяется наличием сложно организованного поддерживающего аппарата, к которому, в частности, относятся телофрагма и мезофрагма. Кальций выделяется из саркоплазматической сети, элементы которой оплетают каждую миофибриллу, после поступления сигнала с сарколеммы по Т-трубочкам (совокупность этих элементов описывается как саркотубулярная система).
Скелетная мышца как орган состоит из пучков мышечных волокон, связанных воедино системой соединительнотканных компонентов (рис. 90). Снаружи мышцу покрывает эпимизий - тонкий, прочный и гладкий чехол из плотной волокнистой соединительной ткани, отдающий вглубь органа более тонкие соединительнотканные перегородки - перимизий, который окружает пучки мышечных волокон. От перимизия внутрь пучков мышечных волокон отходят тончайшие прослойки рыхлой волокнистой соединительной ткани, окружающие каждое мышечное волокно - эндомизий.
Типы мышечных волокон в скелетной мышце - разновидности мышечных волокон с определенными структурными, биохимическими и функцио нальными различиями. Типирование мышечных волокон производится на препаратах при постановке гистохимических реакций выявления ферментов - например, АТФазы, лактатдегидрогеназы (ЛДГ), сукцинатдегидрогеназы (СДГ) (рис. 91) и др. В обобщенном виде можно условно выделить три основных типа мышечных волокон, между которыми существуют переходные варианты.
Тип I (красные) - медленные, тонические, устойчивые к утомлению, с небольшой силой сокращения, окислительные. Характеризуются малым диаметром, относительно тонкими миофибриллами,
высокой активностью окислительных ферментов (например, СДГ), низкой активностью гликолитических ферментов и миозиновой АТФазы, преобладанием аэробных процессов, высоким содержанием пигмента миоглобина (определяющим их красный цвет), крупных митохондрий и липидных включений, богатым кровоснабжением. Численно преобладают в мышцах, выполняющих длительные тонические нагрузки.
Тип IIВ (белые) - быстрые, тетанические, легко утомляющиеся, с большой силой сокращения, гликолитические. Характеризуются большим диаметром, крупными и сильными миофибриллами, высокой активностью гликолитических ферментов (например, ЛДГ) и АТФазы, низкой активностью окислительных ферментов, преобладанием анаэробных процессов, относительно низким содержанием мелких митохондрий, липидов и миоглобина (определяющим их светлый цвет), значительным количеством гликогена, сравнительно слабым кровоснабжением. Преобладают в мышцах, выполняющих быстрые движения, например, мышцах конечностей.
Тип IIА (промежуточные) - быстрые, устойчивые к утомлению, с большой силой, оксилительно-гликолитические. На препаратах напоминают волокна типа I. В равной степени способны использовать энергию, получаемую путем окислительных и гликолитических реакций. По своим морфологическим и функциональным характеристикам занимают положение, промежуточное между волокнами типа I и IIB.
Скелетные мышцы человека являются смешанными, т. е. содержат волокна различных типов, которые распределены в них мозаично (см. рис. 91).
Сердечная поперечнополосатая (исчерченная) мышечная ткань встречается в мышечной оболочке сердца (миокарде) и устьях связанных с ним крупных сосудов. Основным функциональным свойством сердечной мышечной ткани служит способность к спонтанным ритмическим сокращениям, на активность которых влияют гормоны и нервная система. Эта ткань обеспечивает сокращения сердца, которые поддерживают циркуляцию крови в организме. Источником развития сердечной мышечной ткани служит миоэпикардиальная пластинка висцерального листка спланхнотома (целомическая выстилка в шейной части эмбриона). Клетки этой пластинки (миобласты) активно размножаются и постепенно превращаются в сердечные мышечные клетки - кардиомиоциты (сердечные миоциты). Выстраиваясь в цепочки, кардиомиоциты формируют сложные межклеточные соединения - вставочные диски, связывающие их в сердечные мышечные волокна.
Зрелая сердечная мышечная ткань образована клетками - кардиомиоцитами, связанными друг с другом в области вставочных дисков и образующими трехмерную сеть ветвящихся и анастомозирующих сердечных мышечных волокон (рис. 92).
Кардиомиоциты (сердечные миоциты) - цилиндрические или ветвящиеся клетки, более крупные в желудочках. В предсердиях они обычно имеют неправильную форму и меньшие размеры. Эти клетки содержат одно или два ядра и саркоплазму, покрыты сарколеммой, которая снаружи окружена базальной мембраной. Их ядра - светлые, с преобладанием эухроматина, хорошо заметными ядрышками - занимают в клетке центральное положение. У взрослого человека значительная часть кардиомиоцитов - полиплоидные, более половины - двуядерные. Саркоплазма кардиомиоцитов содержит многочисленные органеллы и включения, в частности, мощный сократительный аппарат, который сильно развит в сократительных (рабочих) кардиомиоцитах (в особенности, в желудочковых). Сократительный аппарат представлен сердечными исчерченными миофибриллами, по строению сходными с миофибриллами волокон скелетной мышечной ткани (см. рис. 94); в совокупности они обусловливают поперечную исчерченность кардиомиоцитов.
Между миофибриллами у полюсов ядра и под сарколеммой располагаются очень многочисленные и крупные митохондрии (см. рис. 93 и 94). Миофибриллы окружены элементами саркоплазматической сети, связанными с Т-трубочками (см. рис. 94). Цитоплазма кардиомиоцитов содержит кислородсвязывающий пигмент миоглобина и скопления энергетических субстратов в виде липидных капель и гранул гликогена (см. рис. 94).
Типы кардиомиоцитов в сердечной мышечной ткани различаются структурными и функциональными признаками, биологической ролью и топографией. Выделяют три основных типа кардиомиоцитов (см. рис. 93):
1)сократительные (рабочие) кардиомиоциты образуют основную часть миокарда и характеризуются мощно развитым сократительным аппаратом, занимающим бjльшую часть их саркоплазмы;
2)проводящие кардиомиоциты обладают способностью к генерации и быстрому проведению электрических импульсов. Они образуют узлы, пучки и волокна проводящей системы сердца и разделяются на несколько подтипов. Характеризуются слабым развитием сократительного аппарата, светлой саркоплазмой и крупными ядрами. В проводящих сердечных волокнах (Пуркинье) эти клетки имеют крупные размеры (см. рис. 93).
3)секреторные (эндокринные) кардиомиоциты располагаются в предсердиях (в особенности, пра-
вом) и характеризуются отростчатой формой и слабым развитием сократительного аппарата. В их саркоплазме вблизи полюсов ядра находятся окруженные мембраной плотные гранулы, содержащие предсердный натриуретический пептид (гормон, вызывающий потерю натрия и воды с мочой, расширение сосудов, снижение артериального давления).
Вставочные диски осуществляют связь кардиомиоцитов друг с другом. Под световым микроскопом они имеют вид поперечных прямых или зигзагообразных полосок, пересекающих сердечное мышечное волокно (см. рис. 92). Под электронным микроскопом определяется сложная организация вставочного диска, представляющего собой комплекс межклеточных соединений нескольких типов (см. рис. 94). В области поперечных (ориентированных перпендикулярно расположению миофибрилл) участков вставочного диска соседние кардиомиоциты образуют многочисленные интердигитации, связанные контактами типа десмосом и адгезивных фасций. Актиновые филаменты прикрепляются к поперечным участкам сарколеммы вставочного диска на уровне линии Z. На сарколемме продольных участков вставочного диска имеются многочисленные щелевые соединения (нексусы), обеспечивающие ионную связь кардиомиоцитов и передачу импульса сокращения.
Гладкая мышечная ткань входит в состав стенки полых (трубчатых) внутренних органов - бронхов, желудка, кишки, матки, маточных труб, мочеточников, мочевого пузыря (висцеральная гладкая мышечная ткань), а также сосудов (васкулярная гладкая мышечная ткань). Гладкая мышечная ткань встречается также в коже, где она образует мышцы, поднимающие волос, в капсулах и трабекулах некоторых органов (селезенка, яичко). Благодаря сократительной активности этой ткани обеспечивается деятельность органов пищеварительного тракта, регуляция дыхания, крово- и лимфотока, выделение мочи, транспорт половых клеток и др. Источником развития гладкой мышечной ткани у эмбриона является мезенхима. Свойствами гладких миоцитов обладают также некоторые клетки, имеющие другое происхождение - миоэпителиальные клетки (видоизмененные сократительные эпителиальные клетки в некоторых железах) и мионейральные клетки радужки глаза (развиваются из нейрального зачатка). Структурно-функциональной единицей гладкой мышечной ткани служит гладкий миоцит (гладкая мышечная клетка).
Гладкие миоциты (гладкие мышечные клетки) - вытянутые клетки преимущественно вере-
теновидной формы, не обладающие поперечной исчерченностью и образующие многочисленные соединения друг с другом (рис. 95-97). Сарколемма каждого гладкого миоцита окружена базальной мембраной, в которую вплетаются тонкие ретикулярные, коллагеновые и эластические волокна. Гладкие миоциты содержат одно удлиненное диплоидное ядро с преобладанием эухроматина и 1-2 ядрышками, расположенное в центральной утолщенной части клетки. В саркоплазме гладких миоцитов умеренно развитые органеллы общего значения располагаются вместе с включениями в конусовидных участках у полюсов ядра. Периферическая ее часть занята сократительным аппаратом - актиновыми и миозиновыми миофиламентами, которые в гладких миоцитах не формируют миофибрилл. Актиновые миофиламенты прикрепляются в саркоплазме к овальным или веретеновидным плотным тельцам (см. рис. 97) - структурам, гомологичным линиям Z в поперечнополосатых тканях; сходные образования, связанные с внутренней поверхностью сарколеммы, называют плотными пластинками.
Сокращение гладких миоцитов обеспечивается взаимодействием миофиламентов и развивается в соответствии с моделью скользящих нитей. Как и в поперечнополосатых мышечных тканях, сокращение гладких миоцитов индуцируется притоком Са 2+ в саркоплазму, который в этих клетках выделяется саркоплазматической сетью и кавеолами - многочисленными колбовидными впячиваниями поверхности сарколеммы. Благодаря выраженной синтетической активности гладкие миоциты продуцируют и выделяют (подобно фибробластам) коллагены, эластин и компоненты аморфного вещества. Они способны также синтезировать и секретировать ряд факторов роста и цитокинов.
Гладкая мышечная ткань в органах обычно представлена пластами, пучками и слоями гладких миоцитов (см. рис. 95), внутри которых клетки связаны интердигитациями, адгезивными и щелевыми соединениями. Расположение гладких миоцитов в пластах таково, что узкая часть одной клетки прилежит к широкой части другой. Это способствует наиболее компактной укладке миоцитов, обеспечению максимальной площади их взаимных контактов и высокой прочности ткани. В связи с описанным расположением гладких мышечных клеток в пласте на поперечных срезах соседствуют сечения миоцитов, разрезанных в широкой части и в области узкого края (см. рис. 95).
МЫШЕЧНЫЕ ТКАНИ
Рис. 87. Скелетная поперечнополосатая мышечная ткань
1 - мышечное волокно: 1.1 - сарколемма, покрытая базальной мембраной, 1.2 - саркоплазма, 1.2.1 - миофибриллы, 1.2.2 - поля миофибрилл (Конгейма); 1.3 - ядра мышечного волокна; 2 - эндомизий; 3 - прослойки рыхлой волокнистой соединительной ткани между пучками мышечных волокон: 3.1 - кровеносные сосуды, 3.2 - жировые клетки

Рис. 88. Скелетное мышечное волокно (схема):
1 - базальная мембрана; 2 - сарколемма; 3 - миосателлитоцит; 4 - ядро миосимпласта; 5 - изотропный диск: 5.1 - телофрагма; 6 - анизотропный диск; 7 - миофибриллы

Рис. 89. Участок миофибриллы волокна скелетной мышечной ткани (саркомер)
Рисунок с ЭМФ
1 - изотропный диск: 1.1 - тонкие (актиновые) миофиламенты, 1.2 - телофрагма; 2 - анизотропный диск: 2.1 - толстые (миозиновые) миофиламенты, 2.2 - мезофрагма, 2.3 - полоса Н; 3 - саркомер

Рис. 90. Скелетная мышца (поперечный срез)
Окраска: гематоксилин-эозин
1 - эпимизий; 2 - перимизий: 2.1 - кровеносные сосуды; 3 - пучки мышечных волокон: 3.1 - мышечные волокна, 3.2 - эндомизий: 3.2.1 - кровеносные сосуды
Рис. 91. Типы мышечных волокон (поперечный срез скелетной мышцы)
Гистохимическая реакция выявления сукцинатдегидрогеназы (СДГ)
1 - волокна I типа (красные волокна) - с высокой активностью СДГ (медленные, окислительные, устойчивые к утомлению); 2 - волокна IIВ типа (белые волокна) - с низкой активностью СДГ (быстрые, гликолитические, утомляемые); 3 - волокна IIА типа (промежуточные волокна) - с умеренной активностью СДГ (быстрые, окислительно-гликолитические, устойчивые к утомлению)

Рис. 92. Сердечная поперечнополосатая мышечная ткань
Окраска: железный гематоксилин
А - продольный срез; Б - поперечный срез:
1 - кардиомиоциты (образуют сердечные мышечные волокна): 1.1 - сарколемма, 1.2 - саркоплазма, 1.2.1 - миофибриллы, 1.3 - ядро; 2 - вставочные диски; 3 - анастомозы между волокнами; 4 - рыхлая волокнистая соединительная ткань: 4.1 - кровеносные сосуды

Рис. 93. Ультраструктурная организация кардиомиоцитов различных типов
Рисунки с ЭМФ
A - сократительный (рабочий) кардиомиоцит желудочка сердца:
1 - базальная мембрана; 2 - сарколемма; 3 - саркоплазма: 3.1 - миофибриллы, 3.2 - митохондрии, 3.3 - липидные капли; 4 - ядро; 5 - вставочный диск.
Б - кардиомиоцит проводящей системы сердца (из субэндокардиальной сети волокон Пуркинье):
1 - базальная мембрана; 2 - сарколемма; 3 - саркоплазма: 3.1 - миофибриллы, 3.2 - митохондрии; 3.3 - гранулы гликогена, 3.4 - промежуточные филаменты; 4 - ядра; 5 - вставочный диск.
В - эндокринный кардиомиоцит из предсердия:
1 - базальная мембрана; 2 - сарколемма; 3 - саркоплазма: 3.1 - миофибриллы, 3.2 - митохондрии, 3.3 - секреторные гранулы; 4 - ядро; 5 - вставочный диск

Рис. 94. Ультраструктурная организация области вставочного диска между соседними кардиомиоцитами
Рисунок с ЭМФ
1 - базальная мембрана; 2 - сарколемма; 3 - саркоплазма: 3.1 - миофибриллы, 3.1.1 - саркомер, 3.1.2 - изотропный диск, 3.1.3 - анизотропный диск, 3.1.4 - светлая полоса Н, 3.1.5 - телофрагма, 3.1.6 - мезофрагма, 3.2 - митохондрии, 3.3 - Т-трубочки, 3.4 - элементы саркоплазматической сети, 3.5 - липидные капли, 3.6 - гранулы гликогена; 4 - вставочный диск: 4.1 - интердигитации, 4.2 - адгезивная фасция, 4.3 - десмосома, 4.4 - щелевое соединение (нексус)

Рис. 95. Гладкая мышечная ткань
Окраска: гематоксилин-эозин
А - продольный срез; Б - поперечный срез:
1 - гладкие миоциты: 1.1 - сарколемма, 1.2 - саркоплазма, 1.3 - ядро; 2 - прослойки рыхлой волокнистой соединительной ткани между пучками гладких миоцитов: 2.1 - кровеносные сосуды

Рис. 96. Изолированные гладкие мышечные клетки
Окраска: гематоксилин
1 - ядро; 2 - саркоплазма; 3 - сарколемма

Рис. 97. Ультраструктурная организация гладкого миоцита (участок клетки)
Рисунок с ЭМФ
1 - сарколемма; 2 - саркоплазма: 2.1 - митохондрии, 2.2 - плотные тельца; 3 - ядро; 4 - базальная мембрана
Растительные и животные организмы различаются не только внешне, но и, конечно, внутренне. Однако самая главная отличительная черта образа жизни - это то, что животные способны активно передвигаться в пространстве. Обеспечивается это благодаря наличию в них особых тканей - мышечных. Их мы и рассмотрим подробнее дальше.
Животные ткани
В организме млекопитающих животных и человека выделяют 4 типа тканей, выстилающих все органы и системы, формирующих кровь и осуществляющих жизненно важные функции.
Совокупное сочетание всех перечисленных видов обеспечивает нормальное строение и функционирование живых существ.
Мышечная ткань: классификация
Особую роль в активной жизнедеятельности человека и животных играет специализированная структура. Ее название - мышечная ткань. Строение и функции ее весьма своеобразны и интересны.
Вообще данная ткань неоднородна и имеет свою классификацию. Следует рассмотреть ее подробнее. Существуют такие разновидности мышечных тканей, как:
- гладкая;
- поперечнополосатая;
- сердечная.
Каждая из них имеет свое место локализации в организме и выполняет строго определенные функции.
Строение клетки мышечной ткани
Все три разновидности мышечных тканей имеют свои особенности строения. Однако можно выделить общие закономерности устройства клетки такой структуры.
Во-первых, она удлиненной формы (иногда достигает 14 см), то есть тянется вдоль всего мышечного органа. Во-вторых, она многоядерная, так как именно в этих клетках наиболее интенсивно протекают процессы синтеза белка, образования и распада молекул АТФ.
Также особенности строения мышечной ткани в том, что ее клетки содержат пучки миофибрилл, сформированных двумя белками - актином и миозином. Именно они обеспечивают главное свойство этой структуры - сократимость. Каждая нитевидная фибрилла включает в себя полосы, в микроскоп видимые как более светлые и темные. Ими являются белковые молекулы, образующие что-то вроде тяжей. Актин формирует светлые, а миозин - темные.

Особенности мышечной ткани любого типа в том, что их клетки (миоциты) образуют целые скопления - пучки волокон, или симпласты. Каждый из них изнутри выстлан целыми скоплениями фибрилл, в то время как сама мельчайшая структура состоит из названных выше белков. Если рассмотреть образно данный механизм строения, то получается, словно матрешка, - меньшее в большем, и так до самых пучков волокон, объединенных рыхлой соединительной тканью в общую структуру - определенный тип мышечной ткани.
Внутренняя среда клетки, то есть протопласт, содержит все те же самые структурные компоненты, что и любая другая в организме. Отличие - в количестве ядер и их ориентации не в центре волокна, а в периферической части. Также в том, что деление происходит не за счет генетического материала ядра, а благодаря особым клеткам, носящим название сателлитов. Они входят в состав оболочки миоцита и активно выполняют функцию регенерации - восстановления целостности ткани.
Свойства мышечных тканей
Как и любые другие структуры, данные разновидности тканей имеют свои особенности не только в строении, но и в выполняемых функциях. Основные свойства мышечных тканей, благодаря которым они могут это делать:
- сокращение;
- возбудимость;
- проводимость;
- лабильность.
Благодаря большому количеству кровеносных сосудов и капилляров, питающих мышцы, они могут быстро воспринимать сигнальные импульсы. Данное свойство называется возбудимостью.
Также особенности строения мышечной ткани позволяют ей быстро реагировать на любые раздражения, посылая ответный импульс в кору головного и спинной мозга. Так проявляется свойство проводимости. Это очень важно, так как способность вовремя отреагировать на угрожающие воздействия (химического, механического, физического характера) - важное условие нормальной безопасной жизнедеятельности любого организма.
Мышечная ткань, строение и функции, которые она выполняет - все это в целом сводится к главному свойству, сократимости. Оно подразумевает произвольное (контролируемое) или непроизвольное (без осознанного управления) уменьшение или увеличение длины миоцита. Происходит это благодаря работе белковых миофибрилл (актиновых и миозиновых нитей). Они могут растягиваться и истончаться почти до невидимости, а затем снова быстро восстанавливать свою структуру.
В этом состоят особенности мышечной ткани любого типа. Так построена работа сердца человека и животных, их сосудов, глазных мышц, вращающих яблоко. Именно данное свойство обеспечивает способность к активному движению, перемещению в пространстве. Что бы сумел сделать человек, если бы его мышцы не могли сокращаться? Ничего. Поднять и опустить руку, подпрыгнуть, присесть, танцевать и бегать, выполнять различные физические упражнения - все это помогают делать только мышцы. А именно миофибриллы актиновой и миозиновой природы, образующие миоциты ткани.

Последнее свойство, о котором необходимо упомянуть, это лабильность. Она подразумевает способность ткани быстро восстанавливаться после возбуждения, приходить в абсолютную работоспособность. Лучше миоцитов это могут делать только аксоны -
Строение мышечных тканей, обладание перечисленными свойствами, - главные причины выполнения ими ряда важнейших функций в организмах животных и человека.
Гладкая ткань
Одна из разновидностей мышечных. Имеет мезенхимное происхождение. Устроена отлично от других. Миоциты небольшие, слегка вытянутые, напоминают утолщенные в центре волокна. Средний размер клетки составляет около 0,5 мм в длину и 10 мкм в диаметре.
Протопласт отличается отсутствием сарколеммы. Ядро одно, а вот митохондрий много. Локализация генетического материала, отделенного от цитоплазмы кариолеммой, - в центре клетки. Плазматическая мембрана устроена достаточно просто, сложных белков и липидов не наблюдается. Рядом с митохондриями и по всей цитоплазме разбросаны миофибрилльные кольца, содержащие актин и миозин в небольших количествах, однако достаточных для сокращения ткани. Эндоплазматическая сеть и комплекс Гольджи несколько упрощены и редуцированы по сравнению с другими клетками.
Гладкая мышечная ткань образована пучками миоцитов (веретенообразных клеток) описанного строения, иннервируется эфферентными и афферентными волокнами. Подчиняется управлению вегетативной нервной системы, то есть сокращается, возбуждается без осознанного контроля организма.
В некоторых органах гладкая мускулатура сформирована благодаря индивидуальным одиночным клеткам с особенной иннервацией. Хотя такое явление достаточно редко. В целом можно выделить два основных типа клеток гладкой мускулатуры:

Первая группа клеток малодифференцированна, содержит множество митохондрий, хорошо выраженный аппарат Гольджи. В цитоплазме явно прослеживаются пучки сократительных миофибрилл и микрофиламентов.
Вторая группа миоцитов специализируется на синтезе полисахаридов и сложных комбинативных высокомолекулярных веществах, из которых в дальнейшем строятся коллаген и эластин. Ими же вырабатывается значительная часть межклеточного вещества.
Места локализации в организме
Гладкая мышечная ткань, строение и функции, которые она выполняет, позволяют ей концентрироваться в разных органах в неодинаковом количестве. Так как иннервация не подчиняется контролю со стороны направленной деятельности человека (его сознания), то и места локализации будут соответствующие. Такие, как:
- стенки кровеносных сосудов и вен;
- большая часть внутренних органов;
- кожа;
- глазное яблоко и прочие структуры.
В связи с этим характер активности гладкой мышечной ткани - быстродействующий низкий.
Выполняемые функции
Строение мышечных тканей накладывает прямой отпечаток на выполняемые ими функции. Так, гладкая мускулатура нужна для следующих операций:

Желчный пузырь, места впадения желудка в кишку, мочевой пузырь, лимфатические и артериальные сосуды, вены и многие другиеорганы - все они способны нормально функционировать только благодаря свойствам гладкой мускулатуры. Управление, еще раз оговоримся, строго автономное.
Поперечно-полосатая мышечная ткань
Рассмотренные выше не подчиняются управлению со стороны сознания человека и не отвечают за его движение. Это прерогатива следующего вида волокон - поперечно-полосатых.
Сначала разберемся, за что им было дано такое название. При рассмотрении в микроскоп можно увидеть, что данные структуры имеют четко выраженную исчерченность поперек определенными тяжами - нитями белка актина и миозина, образующими миофибриллы. Это и послужило причиной для такого названия ткани.
Поперечно-мышечная ткань имеет миоциты, содержащие множество ядер и представляющие собой результат слияния нескольких клеточных структур. Такое явление обозначается терминами "симпласт" или "синцитий". Внешний вид волокон представлен длинными, вытянутыми цилиндрическими клетками, плотно соединенными между собой общим межклеточным веществом. Кстати, существует определенная ткань, которая образует эту среду для сочленения всех миоцитов. Ею обладает и гладкая мышечная. Соединительная ткань - основа которая может быть как плотной, так и рыхлой. Она же формирует целый ряд сухожилий, при помощи которых поперечно-полосатая скелетная мускулатура крепится к костям.

Миоциты рассматриваемой ткани, кроме значительного размера, имеют еще несколько особенностей:
- саркоплазма клеток содержит большое количество хорошо различимых микрофиламентов и миофибрилл (актин и миозин в основе);
- данные структуры объединяются в большие группы - мышечные волокна, которые, в свою очередь, формируют непосредственно скелетные мышцы разных групп;
- имеется множество ядер, хорошо выраженный ретикулюм и аппарат Гольджи;
- хорошо развиты многочисленные митохондрии;
- иннервация осуществляется под контролем соматической нервной системы, то есть осознанно;
- утомляемость волокон высокая, однако и работоспособность тоже;
- лабильность выше среднего уровня, быстрое восстановление после рефракции.
В теле животных и человека поперечнополосатая мускулатура имеет красный цвет. Это объясняется присутствием в волокнах миоглобина - специализированного белка. Каждый миоцит покрыт снаружи практически невидимой прозрачной оболочкой - сарколеммой.
В молодом возрасте животных и человека содержат больше плотной соединительной ткани между миоцитами. С течением времени и старением она заменяется на рыхлую и жировую, поэтому мышцы становятся дряблыми и слабыми. В целом скелетная мускулатура занимает до 75% от общей массы. Именно она составляет мясо животных, птиц, рыб, которое человек употребляет в пищу. Питательная ценность очень высокая из-за большого содержания различных белковых соединений.
Разновидностью поперечно-полосатой мускулатуры, помимо скелетной, является сердечная. Особенности ее строения выражаются в присутствии двух типов клеток: обычных миоцитов и кардиомиоцитов. Обычные имеют такое же строение, как и скелетные. Отвечают за автономное сокращение сердца и его сосудов. А вот кардиомиоциты - особые элементы. В них незначительное количество миофибрилл, а значит, актина и миозина. Это говорит о низкой способности к сокращению. Но их задача не в этом. Главная роль - выполнение функции проведения возбудимости по сердцу, осуществление ритмической автоматии.

Сердечная мышечная ткань формируется за счет многократного ветвления входящих в ее состав миоцитов и последующего объединения в общую структуру этих веточек. Еще одно отличие от поперечно-полосатой скелетной мускулатуры - в том, что сердечные клетки содержат ядра в своей центральной части. Миофибриллярные участки локализованы по периферии.
Какие органы образует?
Вся скелетная мускулатура организма - это поперечно-полосатая мышечная ткань. Таблица, отражающая места локализации данной ткани в организме, приведена ниже.
Значение для организма
Роль, которую исполняет поперечно-полосатая мускулатура, переоценить сложно. Ведь именно она отвечает за самое важное отличительное свойство растений и животных - способность к активному передвижению. Человек может совершать массу самых сложных и простых манипуляций, и все они будут зависеть от работы скелетных мышц. Многие люди занимаются тщательными тренировками своей мускулатуры, добиваются в этом большого успеха благодаря свойствам мышечных тканей.
Рассмотрим, какие еще функции выполняет поперечно-полосатая мускулатура в теле человека и животных.
- Отвечает за сложные мимические сокращения, выражение эмоций, внешние проявления сложных чувств.
- Поддерживает положение тела в пространстве.
- Выполняет функцию защиты органов брюшной полости (от механических воздействий).
- Сердечная мускулатура обеспечивает ритмические сокращения сердца.
- Скелетные мышцы участвуют в актах глотания, формируют голосовые связки.
- Регулируют движения языка.
Таким образом, можно сделать следующий вывод: мышечные ткани - важные структурные элементы любого животного организма, наделяющие его определенными уникальными способностями. Свойства и строение разных типов мускулатуры обеспечивают жизненно необходимые функции. В основе строения любой мышцы лежит миоцит - волокно, образованное из белковых нитей актина и миозина.